研究内容
最新の研究成果は随時、学会などにおいて発表しています。また、多くの成果はすでに、英文論文として発表しています。以下のプロジェクトごとの論文要約から、それぞれの論文ごとの主要データ (一部抜粋) と日本語での説明文章を見ることができます。ここで紹介している論文は、当研究室の学生およびスタッフを中心として行ったものです。詳細なデータは英文論文を参照してください。研究室紹介チラシはこちら。
クリプトクロムに関する研究
クリプトクロムは、DNAの紫外線損傷の修復に関わる光回復酵素を含めた大きな遺伝子ファミリーを形成しています。一部を除きほとんどのクリプトクロムは、FAD (フラビンアデニンジヌクレオチド) を結合しており、青色光を受容すると考えられています。
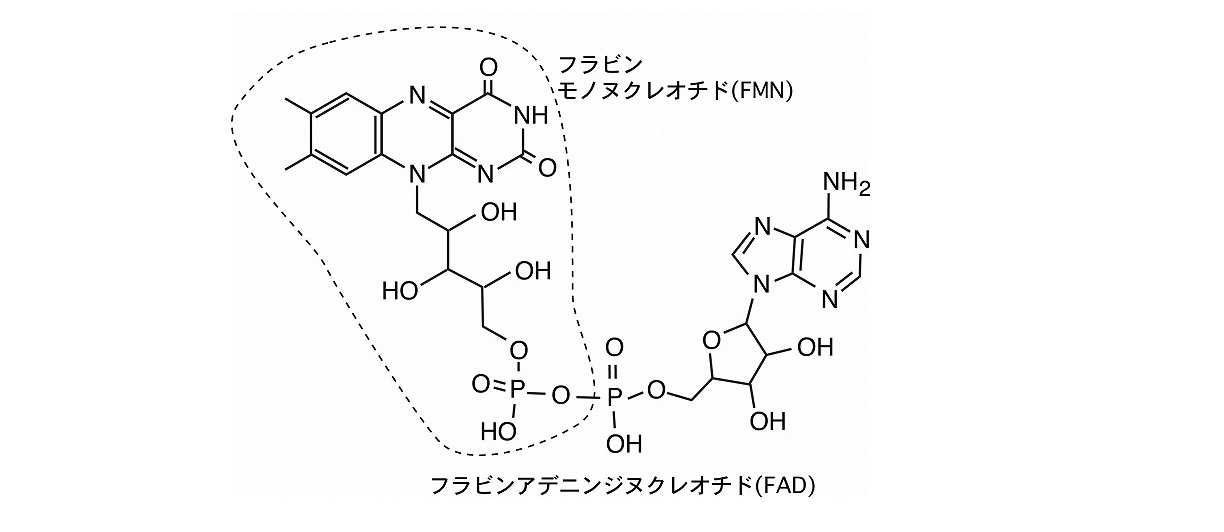
クリプトクロムには複数のサブファミリーがあり、 光による植物の形態形成 や、概日時計の光による同調、あるいは概日時計の発振分子があります。クリプトクロムの研究は主に、これら3つの機能に関して進められていますが、最近では、青色光のエネルギーを利用して地磁気を受容するセンサーとして機能することも明らかになってきました。岡野研究室では、クリプトクロムの光受容機構の研究を中心に、地磁気や微弱光に対する生体の応答機構に関連した研究を進めてきました。
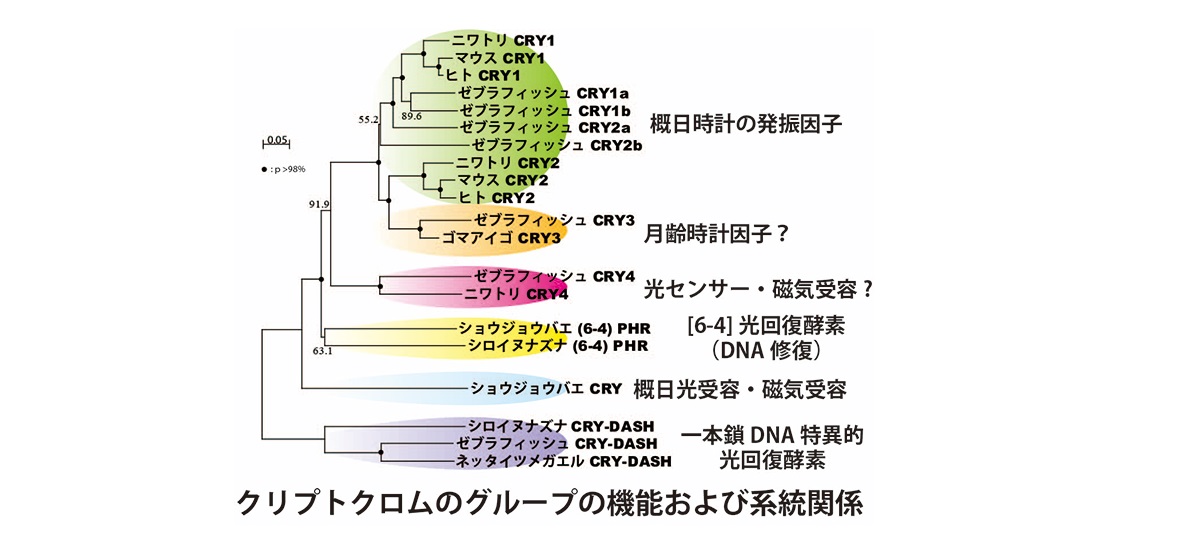
現在のところ、多くの動物が地磁気を感知することが証明されています。しかし、その受容分子の実体や受容のメカニズムは未だよくわかっていません。岡野研では、研究室設立 (2005年) ごろより、脊椎動物のゲノムに存在する機能未知のクリプトクロム遺伝子に着目して研究を始めました。 クリプトクロムは光回復酵素とともに遺伝子ファミリーを形成しており、機能の異なるいくつかのグループに分類されます (図) 。なかでも特に、クリプトクロム4 (CRY4) というファミリーに着目しました。哺乳類にはCRY4は存在しませんが、CRY4をもつ脊椎動物で光依存的な機能を担う2つの可能性を想定しました。1つめは、概日時計の同調に働く光受容体である可能性で、もうひとつは、光エネルギーを利用した地磁気センサーです。現在でもCRY4の生理機能は完全には解明されていませんが、地磁気センサーの可能性が高いと考えています。 まずニワトリ (写真) を材料に、CRY4の解析を行いました。mRNAの発現を各組織でしらべ、抗体を作成して、松果体細胞における発現を細胞レベルで調べました。鳥類の光感受性の概日時計組織である松果体に高い発現が見られました。論文要約PDF : ニワトリ松果体における青色光受容体候補分子クリプトクロム4の同定、発現解析ならびに免疫細胞学的局在解析 [Kubo et al., 2006]、 英語論文はこちら。 多くのニワトリ松果体細胞にCRY4が発現していることがわかりました。しかし、CRY1やCRY2のように概日時計の発振体としての機能は見いだされませんでした。松果体でどのような機能を担っているのかは未だ不明です。 ニワトリを用いた解析では、松果体と並んで、網膜にも高発現することが判りました。地磁気センサーとして機能する際には、網膜に存在する可能性が示唆されており、その予想と見事に一致しました。分子レベルで磁気センサー機能を証明することは難しく、はっきりと示したデータは2018年時点でもありません。脊椎動物のCRYが光を受容するかどうかもわかっておらず、まずはその点を明らかにすることを目指しました。CRY4の解析のために、多くのモノクローナル抗体を研究室内で作製しました。作製した抗体を使ったさまざまな解析の中で、興味深いことに、その中の1つであるC1モノクローナル抗体が、 >明条件よりも暗条件のときに、網膜水溶性画分に存在するCRY4と強く結合することを発見しました。網膜に存在するロドプシンや錐体オプシンは膜タンパク質であり、水溶性画分には含まれません。C1モノクローナル抗体はCRY4のC末端近くと結合することから、網膜に内在するCRY4自身が光を受容してC末端近くの構造が変化し、その結果、抗体との親和性が変化したというモデル (図) を提唱しました。
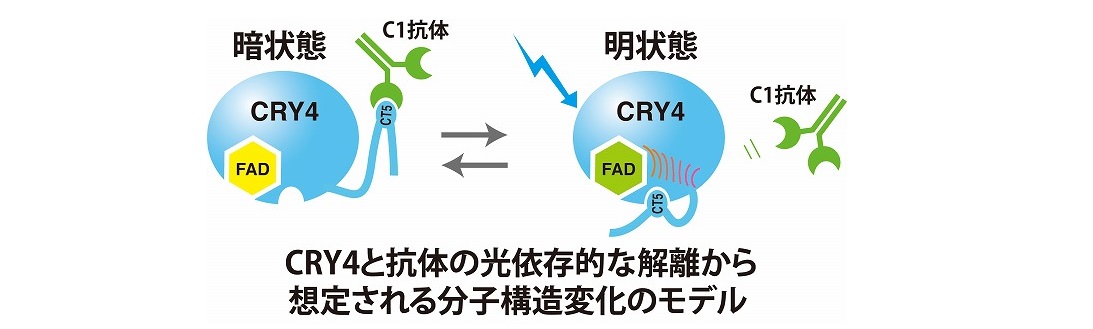
論文要約PDF:ニワトリ網膜におけるクリプトクロム4の光依存的な構造変化 [Watari et al., 2012] 英語論文はこちら ニワトリを用いた研究と並行して、ネッタイツメガエル (Xenopus tropicalis: 写真) を用いたクリプトクロムの解析を行いました。ネッタイツメガエルは、トロピカリスとも呼ばれ、従来から両生類のモデルとして利用されてきたアフリカツメガエル (Xenopus laevis) と近縁関係にあり、アフリカツメガエルにはない多くの利点を備えています。日本国内では特に、広島大学両生類研究センターにおいてバイオリソースとして維持されており、入手も比較的容易です。鳥類だけでなく、爬虫類、両生類、魚類といった進化的に離れた脊椎動物種の同一遺伝子を比較することによって、対象遺伝子の機能の普遍性と進化、さらには環境適応の過程を推察できます。クリプトクロムはおそらく多機能分子と考えられ、単一の動物でも、組織によって機能が異なる可能性があります。そのため、可能な範囲で、複数の動物種で研究を並行して進めることも重要と考えています。 ネッタイツメガエルにもCry4遺伝子がありました。ネッタイツメガエルは、動物個体を用いた行動測定ができ、将来的には遺伝子改変も可能です。そこで、ネッタイツメガエルのクリプトクロムを調べました。ネッタイツメガエルにはCry1, Cry2, Cry4の3つの遺伝子が見つかりました。解析の結果、CRY1, CRY2タンパク質は時計遺伝子Periodの上流プロモータに作用して概日時計遺伝子として働くと推定されました。驚いたことに、Cry1, Cry2のmRNA発現を組織間で比較すると、卵巣においては他の組織に比べて桁違いに非常に高いことがわかりました。 論文要約PDF: ネッタイツメガエルにおいてクリプトクロム遺伝子群は卵巣において高発現している[Kubo, Takeuchi et al., 2010] 英語論文はこちら
一方、ネッタイツメガエルCRY4もニワトリCRY4と同様に、概日時計遺伝子の転写抑制機能は見いだされませんでした。このことは必ずしも、概日時計に無関係であることを直接的に示しているわけではありませんが、核に移行しない点など、概日時計因子としての特徴は備えていません。このように、CRY4はCRY1やCRY2と異なる機能をもつと考えられる一方、mRNA発現を調べると、Cry1, Cry2と同様に、生殖腺 (卵巣および精巣) で非常に高い発現が認められました。このことから、CRYファミリー分子は、概日時計機能とは異なる、生殖腺特異的な機能をもつのではないか、と考えています。 論文要約PDF: ネッタイツメガエル卵巣に発現するクリプトクロム4の同定と性状解析[Takeuchi, Kubo et al., 2014] 英語論文はこちら クリプトクロムの機能を詳細に調べるには、タンパク質を大量に得る必要があります。たとえば1回の可視分光解析には、少なくとも数十μgのタンパク質が必要ですが、比較的タンパク質量が多いニワトリの網膜でも、CRY4は1枚あたりで0.2-0.3μg程度しか含まれていません。そこで次に、培養細胞の中でタンパク質を強制的に大量発現することを試みました。大腸菌やいく種類かの真核細胞を試した結果、出芽酵母の中で、培養液1Lあたり数mgという比較的大量のCRY4タンパク質が得られることがわかりました。タンパク質を精製したところ、幸いFADが結合しており、青色の光を吸収して、FADの還元反応が起こりました。この結果から、ニワトリのCRY4は青色光センサー、もしくは光のエネルギーを利用して地磁気を受容する地磁気センサーではないか、と考えられました。 論文要約PDF: 鳥類磁気受容体候補分子であるクリプトクロム4の酵母を用いた発現と光反応サイクルおよび構造変化 [Mitsui et al., 2015] 英語論文はこちら 岡野研では現在、ニワトリCRY4の一部を改変した変異体タンパク質を調製して、光受容に伴うFADや分子構造の変化などを解析しています。また、ゼブラフィッシュなどニワトリ以外の動物のクリプトクロムの性状解析に向けて、発現系を検討しています。 このように、2006年から当研究室ではニワトリCRY4の研究を行い、ニワトリCRY4が地磁気センサーである可能性を示唆するデータを世界に先駆けて蓄積・発表してきました。当初は他にこの分子を研究するグループはほとんどありませんでしたが、地磁気の受容機構の研究を進める複数のグループが参入しはじめており、今後はニワトリCRY4の研究が加速、競争も激しくなると予想しています。我々が発見したニワトリCRY4が、地磁気センサーであるか、未だはっきりとした証明はありませんが、地磁気センサーとして機能するのであれば、我々自身の手で1番にそれを直接示したいと考えて研究を進めています。
動物の地磁気受容に関する行動学的研究

動物個体を用いた行動学的実験からコマドリやウミガメといった渡り鳥や回遊する動物が地磁気を感知することが、知られていました。しかし、これらの生物は、実験室内での遺伝子レベルでの実験には適しません。一方、ゼブラフィッシュ (写真) は、代表的な魚類のモデル生物であり、遺伝子改変技術も確立されており、飼育も容易であるため、多方面から研究が盛んに行われています。そこで私たちは、ゼブラフィッシュに地磁気受容能力があるかどうかを調べました。さまざまな行動実験系を検討した結果、円形の水槽に放流した際に、一定方向 (南北といった双方向性) に遊泳することがわかりました。 論文要約PDF: ゼブラフィッシュはグループごとに双方向性の地磁気依存的定位を示す [Takebe et al., 2012] 英語論文はこちら
月齢認識機構に関する研究
いくつかの種類の動物では、受精の効率を高め、子孫をなるだけ多く残すために、多数の個体が決まった時に集まって産卵する現象が見られ、同調産卵と呼ばれます。ある種のサンゴは1年に一度だけ、真夏の満月の夜に同調産卵することは有名です。熱帯から亜熱帯に生息するアイゴ類では、種によって異なる月齢の日に同調産卵することが知られています。この月齢同調産卵がどのようにしておこるのか、月光を魚が認識しているのか、また、1ヶ月を周期とするような体内時計が関与しているのか、といった疑問を明らかにするために、上弦の月に合わせて同調産卵するゴマアイゴ (Siganus guttatus: 写真) を材料とした研究を始めました。この研究は琉球大学竹村明洋教授との共同研究です。 はじめに、ゴマアイゴの月齢同調に概日時計因子が関与するかどうかを調べるため、クリプトクロム遺伝子を探しました。同定した2種類のクリプトクロム遺伝子 (SgCry1, SgCry3) に関して、脳と卵巣におけるmRNAの発現を詳細に調べました。その結果、SgCry1は顕著な日内変動を示すもののSgCry3は1日を通して発現量はほぼ一定であることがわかりました。これらの発現を異なる月齢で調べて比較したところ、面白いことに、月齢に依存した発現量の変動が見られました。さらに、その変動パターンを元に、1日の日長をもとに季節を知る光周性と似た仕組みで、月光が時計遺伝子の発現へ影響して月齢応答を引き起こすモデルを提唱しました。 論文要約PDF: サンゴ礁に生息する魚類、ゴマアイゴにおける月齢依存的なクリプトクロムの発現と光周期による月齢認識機構 [Fukushiro et al., 2011] 英語論文はこちら

このように、時計遺伝子が月光認識機構の解明の手がかりとなることがわかったので、続いて、Cry3以外の時計遺伝子は関与しているのか、また、Cry3の周期変動は月光によって制御されているのか、CRY3のタンパク質はどこで働いているのかなどを調べました。概日時計発振系においてCRYと相互作用するPERの遺伝子発現を調べたところ、Per4 (=Per1b) も月齢依存的な変動を示すことがわかりました。この変動が月光の影響下にあるかどうかは、アイゴの水槽を2つに分け、片方だけに毎晩カバーをかけて月光を遮る「月光遮断実験」で調べました。新月から満月まで毎日月光遮断を行なっても、Cry3とPer4の遺伝子変動パターンは影響を受けませんでした。また、CRY3タンパク質は、鳥類で季節応答に関わるとされている脳深部の領域に発現していることも判明しました。これらの結果から、CRY3は月光の刺激で発現が変動するのではなく、少なくとも1月周期前の月光を感じ取って月齢をカウントする内的なタイマー因子であることが示唆されました。 論文要約PDF: 月齢応答性のゴマアイゴにおけるCry3およびPer4の脳深部発現および月光非依存的な変動は月齢時計発振体機能を暗示する [Toda et al., 2014] 英語論文はこちら ここで、新たな疑問が生じました。CRY3は1サイクル前の月光の刺激をもとに振動しているのか、もしくは、もっと長期の内的な概月時計の因子として自律的に約1ヶ月の周期で振動しているのか。近年、海産の環形動物 (ゴカイの1種) において、約1ヶ月の周期で振動する概月時計が存在することが示され、概月時計の仕組みに注目が集まりはじめています。 そこで次に、アイゴのCry3 mRNA振動が、概月時計によるものであれば、数サイクル前からの月光シグナルによって振動していると考えられるため、まるまる2月周期にわたって月光遮断を行い、2周期目に遺伝子振動が観測されるかどうかを調べました。その結果、月光遮断2周期目ではCry3はもはや正常な月光周期下で飼育したゴマアイゴと同じ振動パターンは示しませんでした。このことから、Cry3は1周期前の月光によってリセットされ、次の上弦へ向けて1?2周期の間振動する、いわば砂時計型のタイマー因子であると推定されました。他のCryファミリー分子を含め、Per, Clock, Bmalといった主要な時計遺伝子を多数同時に調べた結果、月光遮断によって、概日時計の振動が乱されていることがわかりました。さらに、間脳から司令をうけて生殖腺刺激ホルモンを分泌する下垂体での遺伝子発現も調べたところ間脳とは異なる発現振動パターンを示すことも判明しました。これらのことは、月齢認識には概日時計が大きく関わっていることを示しており、光周応答に似た仕組みの存在を裏付ける結果といえます。 論文要約PDF: 月齢に同調して産卵するゴマアイゴでは、月光が時計遺伝子発現の月齢依存性および正常な振動を制御している [Takeuchi et al., 2018] 英語論文はこちら
培養細胞の光応答性に関する研究
1998年にUeli Schiblerらは、単離した哺乳類の培養細胞にも概日時計機能が維持されていることを報告しました[Balsaloble et al., 1998]。この発見は、動物の末梢臓器に概日時計機能 (末梢時計) があることと合わせて、概日時計分野の研究を大きく変化させました。また、哺乳類の末梢時計はおそらく光を受容しませんが、昆虫や魚においては、末梢の細胞が直接光を受容して時計が外界と同調することが示されました。 このような背景を受けて当研究室では、ヒト皮膚培養細胞を用いて、本当にヒトの皮膚細胞は光に応答しないのかを検討しました。ヒト皮膚培養細胞にはNB1RGB細胞を用いて、青色光を照射した際の遺伝子変動をマイクロアレイ解析と定量的RT-PCRによって解析しました。その結果、いくつかの遺伝子が強い青色光によって誘導されることが判明し、なかでも、MIC-1と呼ばれるサイトカインはタンパク質レベルでも誘導されました。今回の結果から、ヒトの皮膚において生理的条件でMIC-1が光によって誘導されている可能性がありますが、この実験で用いた光は比較的強いこと、強い短波長光は活性酸素種を生成する反応も引き起こすため、生理的な意義についてはさらなる検討が必要と考えています。 論文要約PDF: ヒト皮膚培養細胞においてマクロファージ抑制性サイトカインであるMIC-1が短波長の光によって誘導される [Akiyama et al., 2009] 英語論文はこちら ヒト皮膚よりも光によく応答する細胞を用いて、外界の光受容と内的な光応答遺伝子や概日時計遺伝子との関連を調べたいと考え、トラフグの眼球由来の培養細胞Fugu Eyeに着目しました。これまでに、ゼブラフィッシュやキンギョの培養細胞が光応答することは知られていましたが、それ以外の魚の細胞については研究がほとんどありませんでした。トラフグ (写真は幼魚) に着目したのは、(1) ゲノムサイズが小さく、ゲノムの配列も決定されていること、(2) フグ類は満月と新月に同調産卵を示す半月周性をもつこと、(3) 完全養殖が最近実現した重要な食糧資源であり効率的かつ安全な栽培漁業の実現に基礎研究が重要であること、などの理由によります。

トラフグのゲノム配列を元に、マイクロアレイを作製し、光応答遺伝子を探索しました。10種類あまりの遺伝子が光応答遺伝子の候補となり、それらの発現解析の結果、光誘導遺伝子を同定することができました。それら遺伝子の光応答性を、概日時計遺伝子と合わせて調べたところ、広い波長領域に渡って敏感に光応答することが判明しました。 論文要約PDF: フグ由来のFugu Eye細胞において、光応答遺伝子と概日遺伝子は広い波長領域の光に応答する [Okano et al., 2017]、 英語論文はこちら。 現在、Fugu Eyeが月光程度の光に応答することができるかを調べています。今後はさらに、月光の変動に応じて、満月や新月を認識するような機構が存在するのかについて調べてゆく予定です。
深海魚の光応答性に関する研究
光環境への適応と進化を研究・考察する上で、深海は魅力的な環境です。深海には、極めて微弱な光しか存在しないため、深海に棲む生物には、眼が退化しているものや、逆に眼が巨大に発達したものがいることは有名です。私たちは、深海のなかでも、熱水噴出孔という特殊環境に適応した、バラビクニンという魚を用いた研究を行いました。熱水噴出孔は通常の深海と異なり、周囲が深くなっており海洋中に孤立して存在する点でも、環境への適応や特異的な進化が期待できます。バラビクニンははっきりとした眼を持っており水深1500m付近に棲んでいます。この研究は、海洋研究開発機構 (JAMSTEC) との共同研究で、沖縄の鳩間海丘の熱水噴出孔でサンプリングをさせていただく機会に恵まれて行なったものです。 バラビクニンのロドプシンの構造を調べ、深海の光環境に適応したアミノ酸置換を起こしていることがわかりました。また、眼球から抽出したロドプシンおよび培養細胞でcDNAを発現させて再構成したロドプシンの分光学的解析を行い、光吸収特性の点でも深海環境に適応していることを明らかにしました。深海環境に棲む生物の多くは、眼が退化しているか、もしくは、浅海との往復の際に眼を利用すると考えられます。一方、熱水噴出孔は、極めて狭いポイントのため、バラビクニンが、熱水噴出孔と浅海の間を往復するとは考えにくく、水深1500mにはほとんど光が到達しないため、なぜ青色光を受容するように進化しているのか興味深い結果です。おそらく深海に適応した発光生物が出す青色の光を検出するのに眼を利用していると想像しています。 論文要約PDF: 暗黒の熱い深海にあるロドプシン:深海熱水噴出孔付近に生息するコンニャク魚バラビクニンがもつロドプシンの分子解析 [Sakata et al., 2015] 英語論文はこちら